Covid-19, de l’insouciance aux interrogations. Un tour d’horizon sur les vaccins, les variants et l’origine du SARS-CoV-2
Modifié le :
Article rédigé le 3 juin 2021 par
DOMINIQUE MORELLO, biologiste moléculaire, retraitée, CNRS
HENRI CAP, zoologue, Muséum d’Histoire Naturelle de Toulouse
Plus d’un an après son apparition, le virus SARS-CoV-2 s’est répandu sur notre planète et y fait toujours des ravages. En un an, les connaissances que nous en avons se sont considérablement étoffées et des mesures prophylactiques, variables d’un pays à l’autre, ont permis de limiter la propagation du virus et de sauver des vies. Mais, au moment où la vaccination se généralise dans les pays du monde entier, il subsiste de nombreuses incertitudes quant à l’évolution du virus (mutations, propagation) et son émergence. Cet article vient compléter les 3 articles déjà parus dans Parlons Science. Il fait un état des lieux des variants et des vaccins disponibles à ce jour et se penche sur l’origine du virus SARS-CoV-2 qu’il parait indispensable d’élucider afin d’éviter de futures crises sanitaires mondiales.
Introduction
L’épidémie due au nouveau coronavirus SARS-CoV-2 a officiellement démarré à Wuhan en République populaire de Chine à la fin de l’année 2019. Elle s’est alors répandue sur le reste de la planète à l’hiver et au printemps 2020. En un an, le virus a touché près de 154 millions de personnes dans le monde et en a tué un peu plus de 3,2 millions, les États-Unis étant le pays le plus endeuillé avec 578 000 morts, suivis du Brésil (409 000 morts), de l’Inde (222 000 morts) et du Mexique (218 000 morts) (chiffres arrondis du 4 mai 2021). En France, plus de 5 millions de cas ont été recensés et nous avons récemment dépassé 105 000 morts déclarées. Sauf exceptions notables (Corée du Sud, Nouvelle-Zélande, …), les mesures sanitaires prises dans la plupart des pays n’ont pas permis de faire disparaître l’épidémie. L’espoir repose désormais sur la vaccination qui devrait, à terme, favoriser une immunité collective et offrir la perspective de « vivre avec » ce virus durablement sans mettre en péril les services hospitaliers et sans bouleverser l’économie.
Nous ne reviendrons pas sur la systématique du SARS-CoV-2 et ses relations de parenté avec les autres coronavirus connus ni sur les différentes étapes de son cycle de vie, ses modes de transmission et les symptômes qu’il provoque puisque ces données sont développées dans un article précédent. Nous consacrerons cet article aux particularités génétiques du SARS-CoV-2 et à son évolution, aux différents vaccins qui sont disponibles ou en préparation et proposerons quelques pistes de réflexion pour évaluer les différentes hypothèses sur l’origine de ce virus.
Particularités génétiques du SARS-CoV-2
Le virus SARS-CoV-2 fait partie des coronavirus ou Coronaviridés, une famille de virus qui parasitent les oiseaux et les mammifères depuis des millions d’années. Parmi ces derniers, les chiroptères ou chauve-souris sont particulièrement concernés puisque sur les 1200 espèces que compte l’ordre, 500 sont porteuses de coronavirus (Sallard et al. 2020). Plus de 500 souches de coronavirus différentes y ont été décrites et certaines espèces en hébergent simultanément plus d’une douzaine (Cyranoski et al. 2020)!
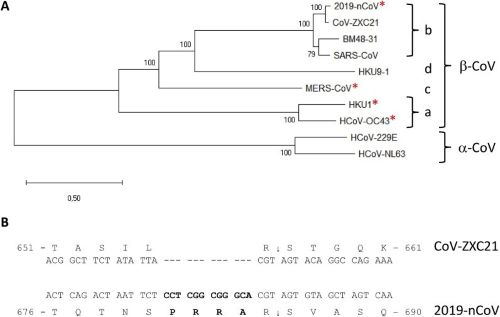
Au sein des coronavirus, sept sont spécifiques à l’espèce humaine (229E, NL63, OC43, HKU1, MERS-CoV, SARS-CoV-1 et SARS-CoV-2). Si les quatre premiers n’engendrent que de simples rhumes, les trois autres ont entraîné des syndromes respiratoires graves. Tous appartiennent à la sous-famille des Orthocoronavirinae. Parmi ces derniers, le SARS-CoV-2 est classé dans le genre Betacoronavirus (B-Cov) (Figure 1) et le sous-genre Sarbecovirus dont les hôtes naturels sont les chauves-souris (Hassanin 2020).
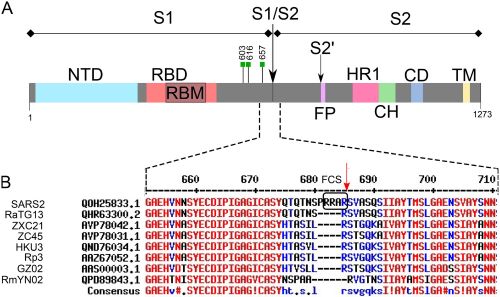
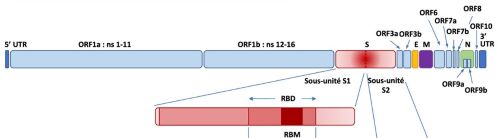
Une des particularités du SARS-CoV-2 est que la protéine S (spike) par laquelle il s’attache au récepteur ACE2 de la cellule hôte porte un site de coupure par la furine. Grace à ce clivage, le virus pénètre très efficacement dans les cellules qu’il infecte. La protéine S est constituée du domaine S1 responsable de la liaison du virus au récepteur (RBD, Receptor Binging Domain) et du domaine S2 responsable de la fusion de l’enveloppe virale avec la membrane cellulaire (Bonnin 2018). Cette étape de fusion nécessite au préalable la coupure de la très grosse protéine S, coupure réalisée par plusieurs enzymes de la cellule hôte, dont la furine qui clive entre les deux domaines S1/S2 et TMPRSS2 (une sérine protéase membranaire) en S’2 (Figure 2).
La furine est exprimée de manière quasi-ubiquitaire par l’ensemble des cellules, quoiqu’à un niveau variable d’un type cellulaire à l’autre, expliquant la grande diversité, ainsi que la gravité des symptômes observés dans les formes sévères de Covid-19 (Coutard et al. 2020). Étrangement, ce site de clivage n’est présent chez aucun autre Betacoronavirus (B-CoV) de la lignée b que le SARS-CoV-2, mais se retrouve, avec une séquence d’acides aminés différente, chez d’autres Betacoronavirus comme MERS-CoV, HKU1 ou OC43 et chez d’autres virus, comme le VIH (Sida), Ebola, la Dengue ou Influenza A (grippe) (voir les virus avec une * dans la Figure 1 ; alignement des séquences dans la figure 2 et Cyranoski 2020).
Une séquence qui bouge : les variants
Le patrimoine génétique (ARN) du virus SARS-CoV-2 a été séquencé à peine quelques semaines après son émergence officielle dans l’est de la Chine à Wuhan en décembre 2019 (Zhou et al. 2020). Fin décembre 2019, les premières séquences étaient disponibles, toutes issues de Wuhan (Chine). À partir de mi-janvier, le séquençage a démarré hors de la Chine. D’abord lent, il s’est considérablement accéléré en avril 2020 avec le confinement massif, quasi-mondial. Plusieurs milliers de nouvelles séquences ont été déposées quotidiennement au printemps sur le site du GISAID (Global Initiative on Sharing Avian Influenza Data, www.gisaid.org/), qui recueille et partage les séquences du monde entier. Aujourd’hui, il héberge plusieurs dizaines de milliers de séquences. Outre son intérêt pour suivre géographiquement la propagation de l’épidémie et comprendre son émergence, le séquençage est indispensable pour suivre les variants portant les mutations déjà identifiées, qui sont parfois non détectables avec les tests PCR, détecter de nouvelles mutations et ainsi comprendre comment le virus s’adapte encore mieux à l’hôte humain et les conséquences des mutations sur sa propagation et sa virulence.
Avec son taux de substitution compris entre 8 10-4 et 8,1 10-3 nucléotide/génome/an, le SARS-CoV-2 évolue presqu’aussi vite que le virus de la grippe (5,2 10-3). Les mutations les plus connues concernent celles qui touchent la protéine S mais on en retrouve sur de nombreuses autres séquences codantes, (par exemple ORF8 et ORF9) (Figure 3).
En ce qui concerne la protéine S, sur les 1273 codons (acides aminés) qu’elle comporte, 28 mutations avaient été recensées fin mars. 11 d’entre elles sont localisées sur la partie N terminale (les mutations des codons 18, 69, 70, 80, 138, 144, 215, 222, 241, 242 et 243). C’est le fragment le plus instable de la protéine S ; 6 mutations affectent le RBD, le domaine de liaison au récepteur (417, 439, 452, 477, 484 et 501) et 4 se trouvent sur les domaines SD1 et SD2 (570, 614, 677 et 681) (Figure 4).
La plus populaire est probablement la substitution D614G dans laquelle une asparagine (D) remplace une glycine (G) au codon 614. Elle apparait d’abord en Italie à la fin janvier 2020, puis se propage rapidement dans le reste de l’Europe et devient dominante en avril, non seulement en Europe mais aussi au Canada, en Australie et une partie des Etats-Unis (Korber et al, 2020 ; Leug et al, 2020). Il semblerait que cette mutation favorise un changement de conformation et améliore la coupure de la protéine S par la furine. Elle augmente la transmissibilité du virus mais n’affecte pas la réponse aux vaccins (voir plus loin).
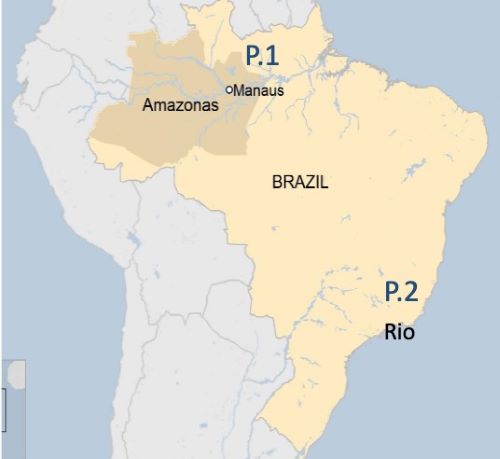
La substitution N501Y est aussi bien connue. Elle est localisée dans le domaine de liaison au récepteur ACE2 (RBD) dont elle modifie le contact. Cette mutation serait responsable de l’augmentation de la contagiosité observée outre-Manche. Les porteurs d’un tel mutant encore appelé variant anglais, ou B1.1.7 – et maintenant renommé le SARS-CoV-2 VOC 202012/01 (Variant of Concern, year 2020, month 12, variant 01) – ont une charge virale importante, ce qui augmente la probabilité de sa propagation. En quelques semaines, ce mutant se propage depuis le Sud-Est de l’Angleterre jusqu’à Londres, ses alentours et bientôt au-delà du Channel où il était présent à la fin du mois de mars dans 114 pays. Ce variant anglais comporte d’autres mutations. Par exemple la délétion de deux acides aminés en position 69 et 70 qui affecte le résultat des tests PCR ciblant le gène S. C’est d’ailleurs une incohérence dans de tels résultats PCR qui a alerté le centre national de référence (CNR) des virus des infections respiratoires de Lyon et lui a permis de détecter le premier variant anglais du SARS-Cov-2 sur le territoire français le 30 décembre 2020. Mais à côté de cette mutation emblématique, une vingtaine d’autres modifications (mutations ou délétions) ont été décrites depuis décembre 2020 sur ce variant anglais qui continue de muter (Figure 4). Certains prélèvements indiquent qu’il porte aussi des changements sur le 484e acide aminé, des mutations qui correspondent à celles que l’on trouve sur les variants « sud-africains » (B.1.351 ou 501Y.V2), apparus depuis octobre 2020, ainsi que du 501Y.V3 (ou P1 et B.1.1.28), puis B.1.1.28.2 ou P.2, dits « brésiliens », apparus en décembre 2020 au Brésil (Figure 5) et dont la prévalence atteint déjà 70 % dans certaines villes.
Figure 5. Répartition géographique des deux mutants brésiliens, P1 et P2.
Fin mars 2021, le variant B1.351 s’est propagé dans 67 pays et le variant P1 dans 36, ces chiffres étant sûrement sous-estimés du fait de l’absence de séquençage dans de nombreux pays.
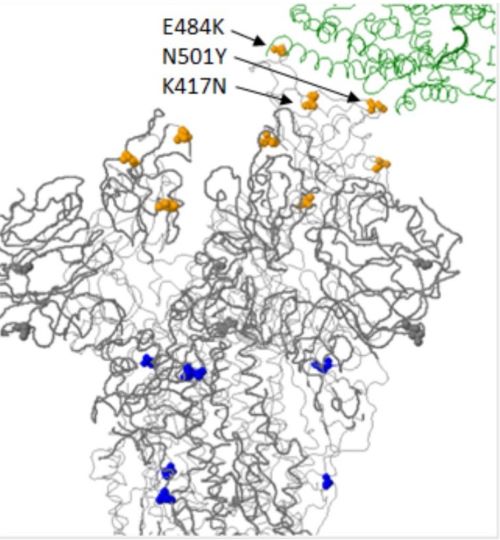
La concomitance des deux mutations N501Y et E484K pourrait donner un avantage au virus, la mutation en position 484 permettant de « doper » celle en 501 et d’améliorer la liaison du virus sur le récepteur cellulaire humain ACE2 (Figure 6). Ces mutations, en particulier sur le 484e acide aminé entrainent la réduction de la fixation des anticorps sur le RBD et de la capacité de neutralisation chez des sujets vaccinés. La mutation E484K pourrait ainsi favoriser la résistance du virus aux anticorps développés dans des sujets déjà contaminés ou vaccinés.
Figure 6. Localisation dans l’espace des 3 mutations sur les acides aminés 417, 484 501 et qui augmentent l’interaction de la protéine S avec le récepteur cellulaire ACE2 (en vert).
Des foyers de nouveaux variants sont détectés maintenant de manière très régulière, par exemple le variant breton 20C/H655Y apparu à la mi-mars en France qui est plus difficile à détecter par un test PCR classique, ou encore le B.1.526 repéré à New-York ou le B.1.429 identifié en Californie. Le variant indien B.1.617, apparu le 5 Octobre 2020 en Inde, est maintenant présent dans plus d’une vingtaine de pays, dont la France. Il porte plusieurs mutations (plus de 15) dont les 3 substitutions L452R, E484Q et P681R. Les deux premières, situées dans le RBD, entraineraient une meilleure fixation du virus au récepteur ACE2 et diminueraient l’efficacité des anticorps neutralisants ; la troisième située dans le site de coupure de la protéine S par la furine pourrait favoriser l’entrée du virus dans les cellules hôtes et augmenter son infectiosité. On ne sait pas encore si ces mutations sont responsables de la flambée récente de l’épidémie en Inde. Ce variant a récemment (le 10 mai) été qualifié de variant préoccupant (VOC) par l’OMS.
La présence de très nombreux variants confirme la nécessité de recourir au séquençage systématique du génome de manière régulière afin de suivre l’évolution du virus et les conséquences de ses mutations sur sa transmissibilité, son infectiosité et son éventuelle capacité à infecter plus facilement d’autres espèces que l’humain.
Rôle du vison dans l’émergence des variants ?
Plusieurs circonstances favorisent l’émergence de mutations. Citons par exemple le cas de patients immunodéprimés infectés de manière chronique par le SRAS-CoV-2. Une étude menée par des virologues de l’Université de Cambridge montre qu’un traitement prolongé d’un malade avec du plasma de convalescent (préalablement infecté par le SARS-CoV-2) favorise chez le même malade l’émergence de mutations (dont la délétion des acides aminés 69-70 et la substitution D796H). Ces mutations entrainent une susceptibilité réduite du virus aux anticorps neutralisants (Kemp et al. 2021).
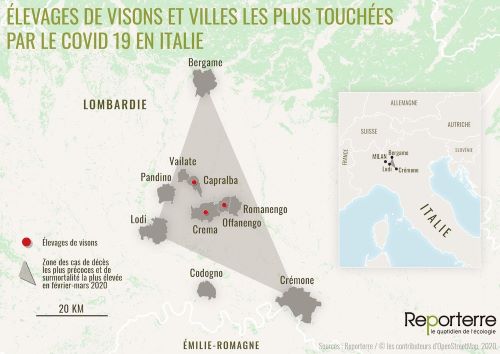
Il est aussi intriguant de constater que de nouveaux variants sont apparus dans des régions d’élevages intensifs de visons, sachant que plus de 30 millions de visons sont répartis dans 2750 élevages intensifs. Les premiers foyers de contamination apparaissent d’abord en Italie début mars 2020, concentrés dans une zone rurale comprise entre les villes de Bergame, Cremone et Lodi (Perrier 2021), où apparait la mutation D614G dès février 2020 (Figure 7).
Figure 7. Relation étroite entre la localisation des élevages de visons et les zones de décès en Italie du Nord, février-mars 2020.
De même en Espagne, à partir du mois de mai, un nouveau variant du virus humain appelé 20A.EU1 (caractérisé principalement par la substitution A222V (Hodcroft et al. 2020)) est localisé en Aragon, près du plus important élevage de visons du pays, d’où il essaimera en juin et durant l’été dans toute l’Europe. Le taux de contamination dans ces élevages est devenu si exponentiel entre mai et juillet qu’il conduira à l’abattage des visons, suspectés d’avoir augmenté la contagiosité du virus, voire d’en être un potentiel hôte intermédiaire (Faure 2020). Le Danemark, premier producteur européen, touché dès le mois de juin a dû abattre 16 millions de visons. Si l’on ne sait pas encore comment les visons ont été contaminés initialement, il est maintenant clair que l’infection en retour, de l’homme par le vison, est effective, comme l’a montré une analyse détaillée portant sur 16 élevages de visons hollandais (Oude Munnink et al. 2021).
Au final, il existe des milliers de variants du SARS-CoV-2. Voir une cartographie complète et actualisée. Cette variabilité génétique a pour conséquence une meilleure adaptabilité du virus à son hôte, une augmentation de l’infectiosité, un échappement aux outils de diagnostic et une résistance aux vaccins de première génération.
Immunité, tests, vaccins et autres traitements
Lorsque les personnes sont infectées, elles développent des anticorps qui persistent une fois le virus éliminé. Il existe une forte hétérogénéité dans la réponse des sujets infectés. On ne connaît pas avec certitude la durée de cette persistance qui est évaluée à 8 mois (Dan et al. 2021), ni le titre minimal, c’est-à-dire la quantité d’anticorps qui confère une immunité protectrice. Les anticorps qui protègent des prochaines infections au SARS-CoV-2 et interrompent le cycle infectieux sont nommésanticorps neutralisants. Ils sont présents, en moyenne 10 à 14 jours après l’apparition des symptômes. Pour évaluer leur capacité de neutralisation, on peut prélever des anticorps d’un patient guéri de la Covid-19 et tester s’ils bloquent l’infection au SARS-CoV-2 chez un animal, le macaque par exemple, ou chez un humain. La preuve absolue de leur capacité à neutraliser reste la prévention d’une réinfection d’un humain par le SARS-CoV-2. C’est l’objectif de la vaccination contre la Covid-19 qui consiste à induire des taux élevés d’anticorps neutralisants sur un grand nombre d’individus ayant été vaccinés. Cependant, on ne sait pas encore si les anticorps produits après vaccination protègeront l’individu immunisé d’une nouvelle réinfection par l’un ou l’autre des variants du SARS-CoV-2. S’il est bien admis que le système immunitaire, en réponse à une infection virale, produit une grande variété d’anticorps ciblant plusieurs protéines et des régions variées d’une même protéine, qu’en est-il du panel d’anticorps synthétisés après une vaccination qui cible simplement la protéine S ? Rien ne garantit qu’ils neutraliseront un variant portant plusieurs mutations sur cette protéine. D’ailleurs, des études récentes font craindre que la mutation E484K portée par les variants sud-africains (B.1.351), brésilien (P1) et récemment découverte sur quelques échantillons de porteurs du variant anglais ne diminue fortement l’action des anticorps neutralisants anti-Covid-19. Les variants porteurs de cette mutation pourraient ainsi échapper aux anticorps, remettant partiellement en cause la stratégie vaccinale actuelle. Citons par exemple l’étude en pré-revue du Professeur Gupta et collaborateurs à Cambridge. Les chercheurs ont fabriqué deux versions d’un pseudovirus portant 8 mutations du variant anglais, l’une avec la mutation E484K l’autre sans celle-ci. Ces pseudovirus ont été mis en contact avec les plasmas de personnes (de 29 à 89 ans) qui avaient reçu 3 semaines auparavant une première dose du vaccin Pfizer-BioNtech. L’étude, qui demande à être répétée avec un plus grand nombre d’échantillons, montre que les personnes de plus de 80 ans ayant reçu une seule dose de vaccin ne sont pas protégées et qu’il faut dix fois plus d’anticorps pour neutraliser le pseudovirus avec la mutation E484K que sans elle (Kemp et al. 2021). Cette approche ne permet cependant pas d’évaluer la contribution de l’immunité cellulaire qui intervient naturellement suite à une infection, mais elle apporte des données indispensables à la mise au point de nouveaux vaccins adaptés aux mutations.
Vaccins de première génération
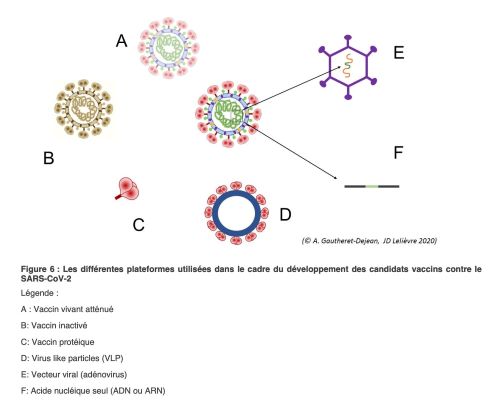
Dans son journal daté du 10 novembre 2020, le CNRS rapporte que 180 candidats-vaccins contre le Covid-19 sont en cours de développement et plus de 40 sont en phase clinique chez l’humain – dont les vaccins des sociétés Pfizer, Moderna ou encore AstraZeneca. De quoi s’y perdre … Sans entrer dans tous les détails, nous allons aborder les caractéristiques principales de ceux qui ont reçu une autorisation de mise sur le marché ou qui sont en phase de l’être. La plupart cible la production d’anticorps contre la protéine Spike qui tapisse la surface du virus, mais les stratégies sont nombreuses (Figure 8). Voir une étude plus complète.
Les deux premiers vaccins disponibles, Moderna et Pfizer/BioNtech sont basés sur une nouvelle technique : elle consiste à injecter un ARN codant la protéine S pleine taille que les cellules du sujet immunisé vont traduire directement en protéine S. La fabrication de cette protéine étrangère à notre corps entrainera petit à petit, à partir d’une dizaine de jours, la production d’anticorps variés reconnaissant différentes régions de cette protéine. Comme l’ARN est un acide nucléique fragile, il est enrobé dans une sorte de nano particule lipidique qui lui permet d’arriver intact dans nos cellules. Ces vaccins nécessitent deux injections successives à environ un moins d’intervalle, la deuxième injection stimulant la production d’anticorps à partir de cellules mémoire générées après la première injection. L’efficacité de ces vaccins est très importante : près de 95%. Avec le recul possible, on sait que des anticorps neutralisants persistent 3 mois après la deuxième injection intra-musculaire du vaccin. Le problème de ces vaccins est leur fragilité : ils nécessitent une conservation à -80°C et une utilisation rapide après décongélation, une logistique incompatible avec leur distribution dans des pays en voie de développement. Leur avantage indéniable est qu’ils sont faciles à mettre en œuvre ; de nouveaux ARN peuvent être synthétisés rapidement (6 semaines), un atout majeur s’il s’avère nécessaire de produire des vaccins contre les variants de SARS-CoV-2 qui pourraient échapper au système immunitaire.
Vaccins génétiques à ADN utilisant un vecteur viral recombinant (Figure 8E)
C’est la stratégie utilisée pour produire de nombreux vaccins (Spoutnik V de l’institut russe Gamaleya, AstraZeneca et l’université d’Oxford, CanSinoBio en Chine, et celui de l’américain par Johnson & Johnson/Janssen). Elle consiste à choisir un vecteur viral inoffensif pour l’humain, ou qui ne peut pas se répliquer car certaines séquences virales ont été retirées, et lui insérer la séquence nucléotidique permettant la production de la protéine Spike. Les virus ainsi recombinés sont injectés et la protéine Spike est synthétisée dans les cellules de la personne vaccinée comme le serait n’importe laquelle des protéines du vecteur viral. Dans tous ces vaccins, le vecteur viral est un virus à ADN responsables de rhumes, l’Adenovirus, mais plusieurs souches d’Adénovirus sont utilisées (Adénovirus humains Adeno 5 et 26 pour Spoutnik V, Adeno 26 pour Johnson & Johnson et Adenovirus de chimpanzé ChAdOx1 pour AstraZeneka). Dans cette stratégie, l’ARN codant Spike est d’abord converti (réverse transcrit) en ADN qui est ensuite intégré dans le vecteur viral à ADN.
Le vaccin AstraZeneca a une efficacité moindre que celle des deux vaccins à ARN (65-90%) mais il ne nécessite pas de protection particulière pour le stockage et peut être conservé plusieurs mois à température ambiante. Après avoir été réservé aux personnes de moins de 65 ans, car les premiers essais cliniques n’avaient pas suffisamment inclus de personnes âgées (>65 ans), son utilisation en France est restreinte aux plus de 55 ans, du fait de très rares cas de thrombose survenus chez des jeunes. D’après les essais cliniques, le vaccin Johnson & Johnson protège à 85 % des formes graves de la pathologie et à 66 % les formes modérées. Cette efficacité moindre que celle des vaccins à ARN pourrait s’expliquer du fait que ce vaccin a été testé plus tardivement sur des populations dans lesquelles les variants s’étaient répandus, en particulier le variant sud-africain B.1.351. Une seule dose suffit.
Notre recul pour évaluer les effets indésirables de tels vaccins génétiques est nul puisqu’aucun vaccin à usage humain utilisant cette technique n’avait encore été commercialisé. Il est toutefois important de signaler que la probabilité de développer une forme grave de la Covid-19 chez les jeunes de 1 à 30 ans est inférieure à celle de développer une thrombose après injection du vaccin.
Vaccin à base de virus inactivé (Figure 8B)
Près d’une vingtaine de projets vaccinaux sont basés sur cette approche. C’est le cas par exemple du vaccin chinois Coronavac (Sinovac) adjuvanté avec des sels d’aluminium, mis à disposition sur le marché avant que les essais cliniques en phase 3 ne soient réellement commencés, le vaccin indien Covaxin ou celui en cours d’étude clinique par la société nantaise Valneva (VLA2001). Le virus SARS-CoV-2 est inactivé par traitement chimique et injecté. Il est ainsi incapable de se répliquer mais a conservé suffisamment de son intégrité physique pour être reconnu comme étranger par le système immunitaire de la personne vaccinée et permettre la synthèse d’anticorps dirigés contre plusieurs protéines du virus (S, M et N). Cependant, du fait des procédés d’inactivation, le vaccin n’est pas très immunogène, il faut répéter les injections et introduire des adjuvants pour induire une protection efficace. Mais cette technologie a l’avantage de nous rassurer car elle est utilisée pour fabriquer de nombreux vaccins (grippe, hépatite A, poliomyélite) et notre recul quant aux effets indésirables est plus important qu’avec les stratégies innovantes. À ce jour, il n’y a encore aucune publication scientifique sur l’efficacité de ces vaccins. Gao Fu, le plus haut responsable médical de Chine, a affirmé le 10 avril que les vaccins chinois (il y en aurait 5 dont Coronavac et CanSino) n’ont pas un taux de protection très élevé, ce qui est regrettable car la Chine a pourvu 160 pays avec ses vaccins!
Vaccins à protéines recombinantes (Figure 8C)
Ces vaccins ont l’avantage de ne pas contenir de matériel vivant mais seulement une protéine. C’est le cas du projet NVX-CoV2373 développé par la société américaine Novavax avec la synthèse de la protéine Spike dans des cellules d’insecte (SF9) et injection sous forme de nanoparticules. Les essais cliniques (phase 3, à petites échelles) montrent une efficacité de 96,4 % contre la souche originale de SARS-CoV-2, 86,3 % contre le variant britannique et 48,6 % contre le variant sud-africain (source Doctissimo). C’est aussi la stratégie que développe Sanofi-Pasteur/GSK en faisant synthétiser la protéine Spike dans des cellules d’insectes. Dans cet exemple, la protéine est codée par un baculovirus recombinant dont le génome contient la séquence codant la protéine Spike. Des cellules d’insectes cultivées en laboratoire sont infectées par ce baculovirus et produisent une grande quantité de protéine Spike qui est ensuite purifiée. Cette stratégie à l’avantage d’être connue puisqu’elle sert déjà par exemple pour produire un vaccin contre la grippe saisonnière ; son inconvénient est qu’il faut rajouter des adjuvants pour booster la réponse immunitaire.
Autres stratégies vaccinales (Figure 8D)
Citons par exemple, la stratégie originale développée par l’entreprise canadienne Medicago. Ce sont des cellules de plantes qui sont utilisées comme « bioréacteurs » pour fabriquer des particules pseudo-virales (PPV) qui imitent la structure naturelle du virus et déclenchent, une fois injectées, une réaction immunitaire.
En résumé, le tableau brossé est loin d’être complet, mais il montre la course dans la production de vaccins ; les gagnants ne seront pas nécessairement les premiers. Les premiers vaccins sur le marché (vaccins à ARN) l’ont été à une époque où peu de variants circulaient et leur efficacité était remarquablement forte. Les derniers vaccins sont testés et utilisés dans un paysage plus complexe dans lequel de nombreux variants sont à l’œuvre, ce qui pourrait expliquer leur efficacité moindre. Ils ont cependant l’avantage d’une conservation moins problématique et sont moins onéreux. Comme tout médicament, les vaccins ont des effets indésirables. Le tout est de savoir évaluer le rapport bénéfice/risque de chacun non seulement pour éviter leurs potentiels effets secondaires indésirables mais aussi pour lutter efficacement contre le SARS-CoV-2 actuel et ses variants, y compris ceux qui ne manqueront pas de circuler dans un proche avenir!
Vaccins en préparation et autres traitements
La plupart des stratégies vaccinales ciblent la protéine S. Certains vaccins de seconde génération en préparation utilisent des versions mutées de la protéine ; d’autres, comme celui développé par le Français OseImmunotherapeutics (CoVepiT) ciblent uniquement l’immunité cellulaire et visent à générer des lymphocytes T mémoires. Le vaccin CoVepit vise la reconnaissance des protéine S, M et N et plusieurs protéines non structurales. On peut espérer qu’il protège contre différents mutants du SARS-CoV-2, présents ou à venir.
Juste avant l’arrivée des premiers vaccins, la compagnie Eli Lilly a obtenu une autorisation de mise sur le marché de ses anticorps monoclonaux dirigés contre la protéine Spike. Leur injection réduirait de 57 % le risque que des personnes âgées contaminées développent des formes sévères de la Covid-19. Ces anticorps administrés préventivement ou en traitement confineraient l’infection au nez et éviteraient les dommages pulmonaires. Il semble qu’ils soient moins efficaces contre le variant sud-africain. Avec l’arrivée des vaccins, ces anticorps monoclonaux devront trouver leur propre niche ; peut-être les personnes âgées dont le système immunitaire n’est plus assez efficace pour répondre au vaccin anti-SARS-CoV-2 (Cohen, 2021)?
Pour finir, donnons une mention particulière à l’Avigan, un médicament utilisé contre la grippe (Favipiravir, T-705). Ce médicament antiviral qui inhibe sélectivement l’ARN polymérase ARN-dépendante du virus de la grippe semble prometteur pour endiguer l’infection au SARS-CoV-2. Cet analogue de nucléoside (les bases du matériel génétique) agirait en permettant la synthèse de génomes viraux tellement altérés que l’enzyme de réparation du virus, une exonucléase normalement très efficace pour rétablir une copie fidèle du génome, serait incapable de réparer (Shannon et al. 2020).
Origine du SARS-CoV-2
Bien qu’on ait acquis en à peine 16 mois de très nombreuses connaissances sur le SARS-Cov-2, qui ont, entre autres, permis de mieux soigner les malades et développer des vaccins en un temps record, la question de l’origine, la source, du SARS-CoV-2 reste étrangement sans réponse. Et pourtant, elle est fondamentale pour éviter une prochaine pandémie virale. Pour simplifier, deux hypothèses sont actuellement plausibles. La première est une zoonose naturelle : le SARS-CoV-2 proviendrait d’un virus de chauve-souris, réservoir naturel de coronavirus, qui aurait contaminé l’humain, directement ou après passage par un hôte intermédiaire, à la faveur d’un rapprochement entre les hommes et les animaux (déforestation, trafic et marché d’animaux sauvages, élevages intensifs,…). L’autre hypothèse serait qu’un (ou plusieurs) virus de chauve-souris aurai(en)t été collecté(s) dans la nature puis introduit dans un des laboratoires de Wuhan, par exemple celui de virologie (WIV) spécialisé dans l’étude des coronavirus de chauves-souris et situé près du marché de poissons d’Huanan reconnu comme amplificateur de l’épidémie. Il y aurait été délibérément adapté aux cellules humaines et se serait accidentellement échappé, contaminant, au gré des rencontres, animaux et humains. Si cette deuxième hypothèse parait a priori invraisemblable et simplement propre à alimenter la théorie du complot – bien qu’un accident ne puisse être considéré comme une conspiration -, de nombreuses données scientifiques sont incompatibles avec l’hypothèse d’une zoonose naturelle. La mission que l’OMS a récemment réalisée en Chine s’est essentiellement concentrée sur la première hypothèse et n’a pas apporté d’éléments permettant d‘éliminer l’hypothèse d’une origine « artificielle » du virus. À ce jour, le mystère reste entier mais les suspicions vont bon train.
Reprenons brièvement l’hypothèse officielle, celle d’une zoonose avec pour point de départ un coronavirus de chauve-souris. Par définition, s’il n’y a pas d’hôte intermédiaire, ce virus doit être adapté aux cellules humaines, c’est-à-dire que sa protéine S, en particulier son domaine de liaison (RBD) doit reconnaitre le récepteur ACE2 des cellules humaines. C’est bien le cas. Mais de manière surprenante, ce domaine est bien mieux adapté au récepteur ACE2 des cellules humaines qu’à celui des cellules de chauve-souris pour lequel il n’a qu’une faible affinité, alors que les chauves-souris sont censées en être à la source (Qiu et al. 2020, Piplani et al. 2020) ! De plus, aucune mutation majeure n’a été enregistrée au début de l’épidémie en Chine chez les premiers malades, contrairement au SARS-CoV de 2002, comme si, dès le départ, le SARS-CoV-2 était déjà adapté à l’homme (Zhan et al. 2020), ce qui ne peut s’expliquer que par une contamination naturelle bien antérieure à novembre 2020, date officielle des premiers cas de Covid-19.
Pour résoudre ce mystère, partons à la recherche de virus « naturels » de chauve-souris dont le SARS-CoV-2 pourrait provenir. Les plus apparentés sont RaTG13 et RmYN02. RaTG13 a été récolté en 2013 (13) dans du guano de chauve-souris de l’espèce Rhinolophus Affinis (Ra) qui résidaient dans la mine de cuivre abandonnée de Tongguan (Tg), au Mojiang dans le Yunnan, à 1100 km de Wuhan (Ge et al., 2013). C’est là qu’en 2012 se sont rendus 6 hommes missionnés pour nettoyer la mine. Ils ont tous les 6 développé une pneumopathie aux symptômes similaires à ceux des personnes infectées par les SARS, et trois en sont morts (Rahalkar & Bahulikar 2020).
Si globalement la séquence de RaTG13 présente un fort pourcentage d’homologie avec celle du SARS-CoV-2 (96,2%), il reste néanmoins 4% de divergence entre les 2 génomes, ce qui, selon la plupart des virologues, correspond à vingt ans de divergence évolutive, voire plus de quarante ans si l’on tient compte de l’adaptation lente à bas bruit dans un hôte intermédiaire sauvage (Andersen et al. 2020, Boni 2020). De plus, la partie de la protéine S qui se lie au récepteur ACE2 ne présente que 77 % d’homologie avec celle du SARS-CoV-2, compromettant une fixation optimale au récepteur ACE2 humaine (Hassanin 2020 ; Zhang et al. 2020). Enfin, la séquence de RaTG13 ne possède pas de site furine (voir Figure 2).
L’autre virus présentant une forte homologie de séquence avec SARS-CoV-2 (93,3 %) s’appelle RmYN02. Il est hébergé par la chauve-souris malaise en fer à cheval, Rhinolophus malayanus (Rm). Il possède un site qui s’apparente au site de coupure par la furine, mais il n’est pas fonctionnel (Segreto & Deigin 2020, Perrier 2021). De plus, sa protéine S et, en particulier son domaine de liaison au récepteur, ne partagent respectivement que 71,9 % et 61,3 % d’homologie avec les parties correspondantes du SARS-CoV-2, ce qui est trop faible pour permettre une liaison optimale au récepteur ACE2 humain.
Deux études publiées il y a à peine deux mois, en février 2021, poursuivent l’investigation en mettant en évidence deux nouveaux coronavirus collectés au Cambodge en 2010 sur la chauve-souris Rhinolophe de Shamel, Rhinolophus shameli, qui partagerait 92,6 % de son patrimoine génétique avec SARS-CoV-2, ce qui en ferait son troisième plus proche parent (Hassanin 2021). La deuxième étude décrit également un nouveau coronavirus, RacCS203, le quatrième plus proche parent de SARS-CoV-2 (91,5 % d’identité), découvert en 2020 dans une grotte en Thaïlande chez la chauve-souris en Fer à cheval acuminé, Rhinolophus acuminatus (Wacharapluesadee et al. 2021). Ce virus, ainsi que RshSTT182 et RshSTT200 les deux souches du Cambodge, seraient en circulation dans le sud-est asiatique depuis 17 ans, puisque des pangolins testés en Chine en 2003 puis en Thaïlande en 2020 avaient développé des anticorps aux SARS-CoV et SARS-CoV-2 ; leur présence confirmerait que les pangolins auraient pu être infectés par ce type de virus durant toute cette période. Mais, comme le précise cette étude, il est impossible de connaître le rôle exact du pangolin : hôte intermédiaire entre la chauve-souris et l’humain, ou troisième maillon de la chaîne ?
Hôte intermédiaire
Ainsi le passage de virus de la chauve-souris à l’humain ne pourrait s’expliquer sans qu’il n’y ait eu un apport d’autres séquences virales par recombinaison, évènements fréquents chez les Sarbecovirus, qui peuvent se dérouler chez la chauve-souris ou après passage dans un hôte intermédiaire (Boni et al. 2020). C’est d’ailleurs ce scenario qui est favorisé pour expliquer les épidémies de SRAS en 2002-2003 et de MERS en 2012, dans lesquelles un virus provenant de chauve-souris aurait transité par la civette masquée, Paguma larvata, pour le SARS-CoV-1 et le dromadaire, Camelus dromedarius, pour le MERS-CoV (Abdel-Moneim, 2014). Toutefois, des analyses récentes suggèrent que ce ne sont pas les civettes qui auraient contaminé l’homme, mais à l’inverse l’homme qui les aurait contaminées lors de leur captivité (Janies et al. 2008, Wenzel 2020a, 2020b). En effet, la recherche de virus sur plus de 1100 civettes de fermes d’élevage s’est révélée négative, tandis que les 6 civettes testées dans un restaurant de Guangzhou fréquenté par des personnes infectées étaient porteuses d’un virus à 99,8 % identique à celui du SARS-CoV-1 (Hassanin 2020). De la même façon, la contamination du dromadaire, réputé hôte intermédiaire pour le MERS, semble avoir été postérieure ou concomitante à la contamination humaine (Machado et al. 2021).
Dans le cas du SARS-CoV-2, en dépit d’un séquençage intensif (plus de 80 0000 échantillons provenant de très nombreuses espèces sauvages, et d’animaux d’élevage – bétails et volailles), il n’y a à ce jour aucun hôte intermédiaire qui résiste à une analyse minutieuse. Même le pangolin a fait long feu. Deux souches apparentées de coronavirus ont été décrites chez le pangolin malais (Manis javanica) (Lam et al. 2020) dont l’une (Guangdong) présente une forte homologie de séquences (97,4 %) avec le SARS-CoV-2 sur une longueur de 74 acides aminés qui correspond au RBD. Pour le reste la séquence du coronavirus de pangolin est plus éloignée de celle du SARS-CoV-2 (91 % d’homologie) que ne l’est celle de RaTG13 (96,4 %). Aucune des séquences virales de pangolin identifiées jusqu’à présent ne présentent le site de coupure à la furine. Enfin, contrairement aux chauves-souris, le pangolin est très sensible au virus, limitant sa diffusion, et il est très rare, classé en danger critique d’extinction, et solitaire (Hassanin 2020). Enfin, RaTG13 ne peut se fixer sur les récepteurs ACE2 du pangolin, ni de chauve-souris, dont il est censé provenir (Mou et al. 2020). Il parait donc fort peu probable que le pangolin soit l’hôte intermédiaire tant recherché de ces deux coronavirus de chauves-souris.
Le vison à nouveau en scène
Si le pangolin ne peut être l’hôte intermédiaire, quel est-il ? Pourrait-il s’agir d’un animal d’élevage ? Un article très récent du WIV propose que le vison puisse être à l’origine de la diffusion du SARS-CoV-2 en Europe lors de la première et de la seconde vague (Hu et Shi, 2021). Selon ces auteurs, les visons seraient responsables de l’apparition des principaux variants européens du virus (D614G), beaucoup plus transmissibles (de 50 à 75 %) du fait des mutations observées sur leurs protéines S (Leug et al. 2020). Ils citent les déclarations à l’Organisation mondiale de la santé animale (WOAH 2020) de huit pays (Danemark, Espagne, États-Unis d’Amérique, Grèce, Italie, Lituanie, Pays-Bas et Suède) signalant la présence de la Covid-19 dans leurs élevages de visons. Ainsi des animaux d’élevage et non plus sauvages pourraient être à l’origine de la pandémie de Covid-19, d’autant que le coronavirus du vison WD1127 aurait les capacités infectieuses les plus proches du SARS-CoV-2 (Guo et al. 2020). Le vison serait-il une simple victime collatérale de la pandémie de Covid-19, comme les chats ou les furets, l’accélérateur et propagateur actif du virus, ou le chaînon manquant entre la chauve-souris et l’humain ? Des données cruciales sur l’évolution sanitaire des élevages de visons et du personnel qui y travaillait au cours de l’année 2019 ainsi que sur les séquences de leur coronavirus nous manquent pour trancher entre ces hypothèses.
Rappelons que les espèces de chauve-souris porteuses de coronavirus sont nombreuses et certaines, comme la Sérotine commune, Eptesicus serotinus, ou le Grand Rhinolophe, Rhinolophus ferrumequinum habitent des régions où se situent les élevages de visons et de chiens viverrins (Wu et al. 2016, Fan et al. 2019, Faure 2021). Des visons infectés auraient pu ensuite être exportés et répandre le SARS-CoV-2 dans les élevages européens. De par leur promiscuité et leur nombre, ils constitueraient un réservoir à mutations. Les variants porteurs des mutations les plus adaptées auraient pu contaminer, en retour, les humains et ce d’autant plus facilement que la région de liaison à la protéine Spike du récepteur ACE2 est très conservée entre le vison et l’homme (Hayashi et al. 2020), ce qui pourrait favoriser les sauts inter-espèces.
Toutefois, si le vison correspondait à l’espèce intermédiaire tant recherchée, avec une protéine S parfaitement adaptée, l’hécatombe aurait dû être observée dans les élevages de visons bien avant que la pandémie ne se déclare à Wuhan. Aucun rapport officiel n’en fait état. L’élevage de visons chinois a pourtant bien dû subir une catastrophe en 2019 puisqu’on a noté une chute brutale de 55 % de la production de fourrure de vison (Faure 2021). Y aurait-il eu une transmission du virus du vison à l’homme bien avant le démarrage officiel de l’épidémie à Wuhan en décembre 2019, virus qui se serait peu à peu adapté à l’humain avant l’« explosion » de Wuhan ?
À quand remonte le début de l’épidémie ?
Plusieurs arguments mettent en doute la piste du marché de fruits de mer Huanan de Wuhan (Fang 2020), un temps suspecté d’être le point de départ de l’épidémie : le premier cas humain recensé porteur de nCov/SARS-CoV-2 ne semble pas avoir fréquenté ce marché (Huang et al. 2020). On n’y trouve pas de petites chauves-souris porteuses de coronavirus proches du SARS-CoV-2 car la population locale n’en consomme pas. De plus, il semblerait que l’origine de la pandémie n’ait pas eu lieu, comme le prétend la version officielle, à la fin du mois de décembre avec un premier foyer de contaminations localisé au marché de Wuhan (Fang 2020, Perrier 2021), mais remonterait en fait au 17 novembre, selon le quotidien South China Morning Post, qui s’appuie sur des données gouvernementales non publiées. Cette même date est aussi avancée par Andrew Rambaut, de l’Université d’Edinbourg, qui conclut, après analyse des différentes souches et mutations du virus à un ancêtre commun datant du même 17 novembre (Andersen et al. 2020). Enfin, un autre événement jette le trouble : le 18 octobre, plus de deux mois avant le premier cas officiel, ont lieu à Wuhan les jeux mondiaux militaires. Le gouvernement et les médias chinois accusent les Etats-Unis d’avoir amené un virus artificiel avec eux et de l’avoir disséminé dans les équipes de militaires sportifs venant du monde entier, dissémination d’autant plus facile que près de 10 000 participants étaient logés au même endroit. Après leur retour, de nombreux cas de maladies suspectes ont été rapportés au sein de la délégation française et italienne. Sur place à Wuhan, cinq militaires sportifs ont été admis dans des hôpitaux pour des crises de malaria, selon les autorités chinoises.
Des travaux réalisés en France très récemment montrent quant à eux que le SARS-CoV-2 circulait déjà dans notre pays dès le mois de novembre 2019 : les chercheurs ont testé la présence d’anticorps anti-SARS-CoV-2 dans près de 10 000 échantillons sanguins collectés sur des participants de la cohorte épidémiologique « Constance » (Carrat et al. 2021). Ils en ont détecté avec une fréquence d’environ 1 cas sur 1000, et ce de manière sporadique sur tout le territoire. Cette étude conforte d’autres travaux italiens suggérant que les premiers cas d’infections par le nouveau coronavirus remonteraient en Italie à octobre 2019 (Apolone et al. 2020). Ainsi le virus avait déjà bien voyagé loin de la Chine avant l’explosion de l’épidémie à Wuhan fin 2019…
Un laboratoire expert en coronavirus
Le laboratoire de Wuhan (WIV), localisé dans la ville d’où l’épidémie a démarré, est spécialisé dans l’étude des coronavirus de chauve-souris. Déjà en 2011-2012, ce laboratoire part à la recherche de coronavirus de chauve-souris apparentés à SARS-CoV, celui qui a provoqué la première épidémie de SRAS en 2002-2003. Des 117 échantillons de guanos de chauve-souris Rhinopholus sinicus rapportés d’une grotte située à Kunming, dans la province de Yunnan, à 1500 km de Wuhan, 27 contenaient des séquences de coronavirus ; deux séquences complètes ont été obtenues (RsSHC014 et RS3367) présentant une très forte homologie avec le SARS-CoV de l’épidémie 2002-2003. Elles ont été propagées dans de nombreuses cellules en culture, dont des cellules de primates et des cellules humaines, permettant d’isoler pour la première fois en laboratoire une souche virale de coronavirus de chauve-souris (bat-SL-CoV-WIV1) capable d’infecter une batterie de cellules nanties d’un récepteur ACE2, dont des cellules humaines, sans qu’il soit nécessaire de recourir à un hôte intermédiaire (Ge et al. 2013).
Plus tard, en collaboration avec plusieurs équipes internationales, ce laboratoire fabrique des virus SARS-CoV « augmentés » pour élaborer des stratégies thérapeutiques et vaccinales en cas d’émergence « naturelle » de ces virus (Menachery et al, 2015 ; 2016 ; Segreto et Deigin 2020). Par exemple, des virus chimériques hautement pathogènes sur des cellules de poumon humaines ont été construits en insérant la séquence de la protéine S d’un virus de chauve-souris dans une séquence de coronavirus de souris. De telles expériences dites de gain de fonction (GOF) auraient pu aboutir à l’émergence d’un virus de type SARS-CoV-2.
Une publication très récente donne du poids à cette hypothèse en montrant comment une souche de coronavirus peu adaptée aux cellules humaines car elle ne possède pas le site de clivage par la furine devient pathogène (Kaina, 2021). Comme mentionné précédemment, ce site furine est très important pour que le virus pénètre dans les cellules car il permet de cliver la protéine S à la jonction des deux domaines S1 et S2 et favorise ainsi la fusion du virus avec la membrane de la cellule hôte. Il est présent dans la séquence du SARS-CoV-2 mais étrangement absent de celle des deux coronavirus de chauve-souris les plus apparentés, RaTG13 et RmYN02 (Zhou et al. 2020). Il n’est pas non plus présent chez le virus du pangolin. En revanche, il existe chez d’autres coronavirus plus éloignés, comme le MERS (Coutard et al. 2020) ou, d’autres virus que les coronavirus, tels que celui de la grippe, d’Ebola ou du VIH (Cyranoski 2020) (Figure 9).
Figure 9. Comparaison de la séquence de la protéine Spike du SARS-CoV-2 avec celles d’autres virus apparentés (96,2 % d’identité avec la souche Bat-CoV RaTG13 isolée de la chauve-souris en fer à cheval ; 79,5 % d’identité avec SARS-CoV (première épidémie de SARS 2002-2003) et 91 % d’identité avec le virus isolé du pangolin. A. Génome du SARS-CoV-2. B. La protéine S qui une fois liée au récepteur ACE2 est coupée en deux sous-unités S1 et S2 par des protéases cellulaires TMPRSS2 et la furine. C. Le site de clivage de la furine à la fin de la séquence du tétrapeptide Arg-Arg-Ala-Arg est marqué par une flèche (S1/S2). Le site de clivage est spécifique du SARS-CoV-2. D. Une insertion similaire se trouve dans la séquence du MERS-CoV. Les séquences possibles pour les codons thréonine et proline sont écrites en dessous.
Comme le montre Kaina dans son article, les séquences du MERS-CoV et du SARS-CoV-2 autour de ce site sont très similaires non seulement au niveau des acides aminés mais aussi des nucléotides (Figure 9B). Plus précisément, la thréonine en position 678 est codée par le même triplet ACT et la proline (681) par le même triplet CCT. Or, vu la redondance du code génétique, la probabilité de trouver simultanément le même triplet pour la thréonine (qui peut être codée par les 4 triplets ACA, ACG, ACC et ACT) et la proline (qui peut être codée par CCT, CCC, CCA et CCG) est de 1/16 soit 0,06 %… Autant dire très faible et qui favorise l’hypothèse d’une acquisition par recombinaison avec le MERS-CoV plutôt que celle d’une coïncidence. Le scénario de l’émergence du SARS-CoV-2 pourrait alors être le suivant : des cellules de mammifères exprimant le récepteur ACE2 auraient pu être infectées simultanément avec le MERS-CoV et le précurseur du SARS-CoV-2 (RaTG13, RmYN02, WIV1 ou un virus hybride portant un RBD adapté au récepteur ACE2). Au fur et à mesure des passages cellulaires, les variants possédant les mutations qui leur permettent de s’adapter à leur nouvel hôte sont sélectionnés, en particulier ceux dans lesquels le site furine qui augmente son pouvoir infectieux s’est inséré (Wang et al. 2020). Il est aussi possible que de tels passages visant, par sélection naturelle, à faciliter l’adaptation du virus à un nouvel hôte, aient été effectués en parallèle ou simultanément in vivo dans des animaux de laboratoires. Ainsi sont couramment utilisés des furets (cousins mustélidés du vison) ou des souris transgéniques humanisées exprimant à la surface de leurs cellules le récepteur ACE2 humain. Des rats nouveaux-nés ont aussi été infectés par l’équipe de Chanjun Wang avec deux coronavirus « naturels » de chauve-souris Rhinolophus sinicus, ZC45 et ZXC21, découverts entre 2015 et 2017, à Zhoushan dans l’Est de la Chine, province du Zhejiang, (Hu et al. 2018). Il est enfin possible que ce site furine ait été construit de toutes pièces puis inséré artificiellement. Un fort argument dans ce sens vient de l’analyse des séquences codant les deux arginines successives du site furine (PRRA), succession qui ne se rencontre chez aucun autre coronavirus. Elles sont codées par la séquence CGG CGG. Alors que seulement 5 % des arginines du SARS-2-CoV-2 sont codées par un tel codon CGG, il se trouve que c’est un des codons privilégiés pour l’arginine dans les cellules humaines.
Que le virus ainsi amélioré sorte accidentellement des laboratoires ne dépend plus que des conditions sanitaires et de sécurité. Or, comme l’a mentionné Botao Xiao, professeur de biologie moléculaire de l’Université de Chine du Sud ou encore le directeur du laboratoire P4 du WIV Yuan Zhiming, elles ne sont pas idéales ! Pour étayer ce scénario, rappelons que des essais cliniques d’un premier vaccin chinois contre le SARS-CoV-2 ont commencé en mars 2020, une prouesse en termes de délai si l’on n’envisage pas qu’un virus vivant atténué était déjà disponible depuis plusieurs mois. Est-il possible que cette stratégie vaccinale ait été testée auparavant en laboratoire et qu’elle ait malencontreusement abouti à la dissémination du virus (Segreto & Deigin 2020, Perrier 2021) ?
Un autre type d’accident aurait pu se produire lors de la collecte et l’isolement d’un virus de chauve-souris, comme celui de 2012, responsable de la mort de mineurs dans le sud de la Chine, ou bien lors de l’analyse d’un potentiel virus pathogène apparu dans les élevages de visons chinois en 2019, responsable de la perte de la moitié de la production.
Conclusions
À ce stade, les interrogations sont encore malheureusement trop nombreuses pour que l’origine du virus puisse être établie avec certitude. Cependant, les investigations menées jusqu’à présent démontrent l’urgence de limiter les risques naturels de zoonoses. Il est plus que temps d’appliquer les lois sur la protection des espèces sauvages menacées, qui interdisent définitivement le commerce de celles qui en sont à l’origine (Hassanin 2020), de réguler la déforestation et le développement des activités agricoles et économiques à outrance pour ne plus favoriser l’émergence de virus à partir d’espèces qui n’avaient auparavant pas de contact avec l’humain. Il parait aussi urgent de réglementer les conditions d’élevage d’espèces potentiellement hôtes intermédiaires de coronavirus, comme le vison (Sallard et al. 2020). Enfin, il est essentiel de faire la lumière sur l’origine du Covid-19. Un groupe de 26 scientifiques de différentes disciplines et nationalités ont écrit une lettre ouverte le 4 mars à la suite de la parution du rapport de l’OMS après la visite en Chine en janvier 2021, dans laquelle ils décrivaient leurs craintes que l’équipe conjointe du comité international et du gouvernement chinois « n’ait pas le mandat, l’indépendance ou les accès nécessaires pour mener une enquête complète et sans restriction sur toutes les hypothèses concernant les origines du SARS-CoV-2″ (Butler et al. 2021a). Dans une deuxième lettre ouverte datée du 7 avril ces mêmes scientifiques soutiennent la déclaration datée du 30 mars 2021 du Directeur général de l’OMS, le Dr Tedros Adhanom Ghebreyesus selon laquelle « toutes les hypothèses au sujet des origines doivent être examinées, y compris la possibilité d’un accident lié à un laboratoire et que la Chine devrait être plus diligente pour partager données et échantillons biologiques ». Dans cette lettre, plusieurs étapes sont détaillées, en particulier un appel à améliorer la surveillance réglementaire nationale et internationale des laboratoires effectuant des recherches virologiques à haut risque, en mettant l’accent sur « les recherches de gain de fonction préoccupantes » (GOFROC). Une commission internationale, indépendante de l’OMS, devrait pouvoir effectuer des contrôles dans n’importe quel pays, afin de vérifier que les recherches sur les vaccins ou sur l’émergence de nouveaux virus ne puissent mener à une épidémie de type SARS-CoV-3 ou aboutir de façon détournée ou à l’occasion d’un changement de régime, à la création d’armes biologiques (Butler et al. 2021b). Encore plus récemment, une troisième lettre ouverte datée du 30 avril adressée à l’OMS soulève un certain nombre de points qui devraient être examinés en détails concernant le début réel de l’épidémie, l’effacement des bases de données en ligne du WIV, les données sur les virus apparentés à SARS-CoV-2 découverts en 2013, ainsi que sur d’autres dissimulations et conflits d’intérêts avérés (Butler et al. 2021c). Le but de ces trois lettres demandant que la lumière soit faite sur l’origine du Sars-CoV-2 a été atteint : le 14 mai 2021, dix-huit scientifiques renommés, pour la plupart virologues américains, ont écrit une lettre ouverte publiée par la revue Science pour que l’OMS ne se contente pas d’étudier la piste de la zoonose, privilégiée jusque-là, mais élargisse sa recherche sur l’origine du virus à une fuite accidentelle de laboratoire (Bloom et al 2021). Cette demande vient ainsi renforcer le travail du groupe des vingt-six (aussi appelé groupe de Paris ou « outsiders » par ses détracteurs) dont une partie provient du groupe DRASTIC qui réunit des chercheurs internationaux indépendants animés par le désir de mieux connaitre les caractéristiques de ce virus hors-norme et son origine, connaissances indispensables pour se prémunir d’une nouvelle pandémie ou tenter de la maîtriser plus rapidement et efficacement.
Références
Abdel-Moheim, A. 2014 Middle East respiratory syndrome coronavirus (MERS-CoV): evidence and speculations, Arch Virol, 159(7): 1575–1584.
Andersen, K.G., A. Rambaut, W.A. Lipkin, E.C. Holmes & R.F. Garry. 2020. The proximal origin of SARS-CoV-2, Nature medecine, 26 : 450-452.
Apolone, A, E. Montomoli, A. Manenti, M. Boeri, F. Sabia, I. Hyseni, L. Mazzini, D. Martinuzzi, L. Cantone, G. Milanese, S. Sestini, P. Suatoni, A. Marchianò, V. Bollati, G. Sozzi & U. Pastorino. 2020. Unexpected detection of SARS-CoV-2 antibodies in the prepandemic period in Italy. Tumori Journal, 11 novembre 2020 : 1-6.
Bloom, J. D., Y. A. Chan, R. S. Baric , P. J. Bjorkman, S. Cobey, B. E. Deverman, D. N. Fisman, R. Gupta , A. Iwasaki, M. Lipsitch, R. Medzhitov, R. A. Neher, R. Nielsen, N. Patterson, T. Stearns, E. van Nimwegen, M. Worobey & D. A. Relman. 2021. Investigate the origin of COVID19. Science, 372 (6543): 694. 14 mai 2021. DOI: 10.1126/science.abj0016
Boni MF, Lemey P, Jiang X, et al.2020. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible fort he COVOD 19 pandemic. Nat Microbiol. 2020 Nov;5(11):1408-1417. doi: 10.1038/s41564-020-0771-4. Epub 2020 Jul 28. PMID: 32724171
Butler, C. D., B. Canard, H. Cap, Y. A. Chan, J-M. Claverie, F. Colombo, V. Courtier, F. A. de Ribera, E. Decroly, R. de Maistre, G. Demaneuf, R. H. Ebright, A. Goffinet, F. Graner, J. Halloy, M. Leitenberg, F. Lentzos, R. McFarlane, J. Metzl, D. Morello, N. Petrovsky, S. Quay, M. C. Rahalkar, R. Segreto, G. Theißen & J. van Helden. 2021a. Open Letter #1: Reaction to the China-WHO joint press conference on the 9th Feb 2021. Call for a Full and Unrestricted International Forensic Investigation into the Origins of COVID-19. March 4 2021. DOI:10.13140/RG.2.2.26695.83368
Butler, C. D., H. Cap, J-M Claverie, F. Colombo, F. A. de Ribera, R. de Maistre, G. Demaneuf, R. H. Ebright, A. Goffinet, F. Graner, J. Halloy, M. Itoh, H. Kakeya, R.M. Leitenberg, F. Lentzos, J. Metzl, D. Morello, N. Petrovsky, S. Quay, M. C. Rahalkar, R. Segreto, G. Theißen, J. van Helden & R. Wiesendanger. 2021b. Open Letter #2: Reaction to the China-WHO joint study team report. Call for a Full Investigation into the Origins of COVID-19. April 7, 2021. DOI:10.13140/RG.2.2.25018.11206
Butler, C. D., H. Cap, J-M Claverie, F. Colombo, V. Courtier, E. Decroly, R de Maistre, F. A. de Ribera, G. Demaneuf, R. H. Ebright, A. Goffinet, F. Graner, J. Halloy, H. Kakeya, R.M. Leitenberg, F. Lentzos, R. McFarlane, J. Metzl, D. Morello, N. Petrovsky, S. Quay, M. C. Rahalkar, R. Segreto, G. Theißen, J. van Helden & R. Wiesendanger. 2021c. Open Letter #3 to the World Health Organisation and the Members of its Executive Board ahead of the World Health Assembly (May 2021).Open Letter to the World Health Organization and the Members of its Executive Board. April 30, 2021. DOI:10.13140/RG.2.2.18097.51041
Cap, H. & D. Morello. 2020. Covid-19 : de l’insouciance au questionnement sur l’origine de SARS-CoV-2. Bulletin de la Société d’Histoire Naturelle de Toulouse, 156 : sous presse. Version française – Version anglaise
Carrat, F., J. Figoni, J. Henny, JC Desenclos, S. Kab, X. de Lamballerie & M. Zins. 2021. Evidence of early circulation of SARS-CoV-2 in France: findings from the population-based “CONSTANCES” cohort. European Journal of Epidemiology, 6 février 2021, https://doi.org/10.1007/s10654-020-00716-2.
Cohen, J. 2021. Monoclonal antibodies can prevent COVID-19 but successful vaccines complicate their future, Science, 22 janvier 2021, doi:10.1126/science.abg7103
Coutard, B., C. Valle, X. de Lamballerie B. Canard, N. G. Sayda & E. Decroly. 2020. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade, Antiviral Research, 176 : 104742.
Cyranoski, D. 2020. Profile of a killer virus. Nature, 581, 22-26.
Dan, J.M., J. Mateus, Y. Kato, K.M. Hastie, E.D. Yu, C.E. Faliti, A. Grifoni, S.I. Ramirez, S. Haupt, A. Frazier, C. Nakao, V. Rayaprolu, S.A. Rawlings, B. Peters, F. Krammer, V. Simon, E. O. Saphire, D. M. Smith, D. Weiskopf, A. Sette & S. Crotty. 2021. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science, 371 (6529) : eabf4063.
Fan, Y., K. Zhao, Z-L. Shi & P. Zhou. 2019. Bats coronaviruses in China, Viruses, 11(3) : 210.
Faure, Y. 2020. Les élevages de visons ont un rôle dans la pandémie, Journal Reporterre, 10 novembre 2020.
Ge, X.Y, Li, J.L., Yang, X.L. et al. 2013. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature, 503, 535.
Guo, Q., M. Li, C. Wang, P. Wang, Z. Fang, J. Tan, S. Wu, Y. Xiao & H. Zhu. 2020. Host and infectivity prediction of Wuhan 2019 novel coronavirus using deep learning algorithm. BioRxiv. https://doi.org/10.1101/2020.01.21.914044
Hassanin, A. 2020. Cov-19 : Origine naturelle ou anthropique? The conversation. 15 Avril 2020.
Hassanin, A. 2021. Il y a 10 ans, un virus proche du SARS-CoV-2 circulait déjà au Cambodge. The conversation. 1er Février 2021.
Hodcroft, E.B., Zuber, M., Nadeau, S., et al. 2020, Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020, medRxiv, 2020 Nov 27.
Janies D., F. Habib, B. Alexandrov, A. Hill & D. Pol. 2008. Evolution of genomes, host shifts and the geographic spread of SARS-CoV and related coronaviruses. Cladistics, 24 : 111-130.
Kaina, B. 2021. On the Origin of SARS-CoV-2: Did Cell Culture Experiments Lead to Increased Virulence of the Progenitor Virus for Humans? In Vivo, 35, doi:10.21873/invivo.11
Kemp, S.A., D.A Collier, R. Datir, et al. SARS-CoV-2 evolution during treatment of chronic infection. 2021, Nature 592, 277-297,
Korber, B., Fischer, W.M., Gnanakaran et al. Changes in SARS-CoV-2 Spike : evidence that D614G increases infectivity of the Covid-19 virus. 2020. Cell, 182, 812-827
Lau et al. 2017 : Molecular Evolution of MERS Coronavirus: Dromedaries as a Recent Intermediate Host or Long-Time Animal Reservoir? Int. J. Mol. Sci. 2017, 18, 2138.
Leung, K., Y. Pei, G.M. Leung, T. Lam & J. Wu. 2020. Empirical Transmission Advantage Of The D614G Mutant Strain Of SARS-Cov-2. medRxiv 2020.09.22.20199810
Machado, D. J., R. Scott, S. Guirales & D.A. Janies. 2021. Fundamental evolution of all Orthocoronavirinae including three deadly lineages descendent from Chiroptera-hosted coronaviruses: SARS-CoV, MERS-CoV and SARS-CoV-2. Cladistics: 1-28.
Menachery, V. D., B. L. Yount Jr, K. Debbink, S. Agnihothram, L. E. Gralinski, J. A. Plante, R. L. Graham, T. Scobey, X-Y Ge, E. F. Donaldson, S. H Randell, A. Lanzavecchia, W. A. Marasco, Z. Shi & R. Baric. 2015. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nature medicine, 21: 1508-1513.
Menachery, V.D., B.L. Yount, A. C. Sims, K. Debbink, S. S. Agnihothram, Gralinski L.E.,
R. L. Graham, T. Scobey, J. A. Plante, S. R. Royal, J. Swanstrom, T. P. Sheahan, R. J. Pickles, D. Corti, S. H. Randell, A. Lanzavecchia, W. A. Marasco & R. S. Baric. 2016. SARS-like WIV1-CoV poised for human emergence. Proceedings of the National Academy of Sciences,113 : 3048-3053.
Mou, H., B. D. Quinlan, H. Peng, Y. Guo, S. Peng, L. Zhang, M.E. Davies-Gardner, M.R. Gardner, G. Crynen, Z. X. Voo, C.C. Bailey, M.D. Alpert, C. Rader, H. Cohe L. & M. Farzan. 2020. Mutations from bat ACE2 orthologs markedly enhance ACE2-Fc neutralization of SARS-CoV-2. BioRxiv. https://doi.org/10.1101/2020.06.29.178459.
Oude Munnink B.B., R. S. Sikkema, D. F. Nieuwenhuijse, R. J. Molenaar, E. Munger, R. Molenkamp, A. van der Spek, P. Tolsma, A. Rietveld, M. Brouwer, N. Bouwmeester-Vincken, F. Harders, R. Hakze-van der Honing, C. A. Wegdam-Blans, R. J. Bouwstra, C. GeurtsvanKessel, A. A. van der Eijk, F. C. Velkers, L. A. M. Smit, A. Stegeman, W. H. M. van der Poel & M. P. G. Koopmans. 2021. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science, 371 (6525) : 172-177.
Perrier B. 2021. SARS-CoV-2 : Aux origines du mal. Belin, Paris.
Piplani S., P. K. Singh, D. A. Winkler & N. Petrovsky. 2020. N silico comparison of spike protein-ACE2 binding affinities across species; significance for the possible origin of the SARS-CoV-2 virus. arXiv:2005.06199v2.
Qiu, Y., Y.B. Zhao, Q. Wang, J.Y. Li, Z.J. Zhou, C. H. Liao & X.Y. Ge. 2020. Predicting the angiotensin converting enzyme 2 (ACE2) utilizing capability as the receptor of SARS-CoV-2. Microbes and infections, 22 : 221-225.
Rahalkar, M. C. & R. A. Bahulikar. 2020. Lethal Pneumonia Cases in Mojiang Miners (2012) and the Mineshaft Could Provide Important Clues to the Origin of SARS-CoV-2. Front. Public Health 8:581569. doi: 10.3389/fpubh.2020.581569.
Sallard, E., J. Halloy, D. Casane, J. Van Helden & E. Decroly. 2020. Retrouver les origines du SARS-CoV-2 dans les phylogénies de coronavirus. Médecine Sciences 36 : 783–796.
Segreto, R. & Y. Deiguin. 2020. The genetic structure of SARS‐CoV‐2 does not rule out a laboratory origin. BioEssays 202000240, 1-9.
Segreto, R., Y. Deigin, K. McCairn, A. Sousa, D. Sirotkin, K. Sirotkin, J.J. Couey, A. Jones & D. Zhang. 2021. Should we discount the laboratory origin of COVID-19? Environmental Chemistry Lettershttps://doi.org/10.1007/s10311-021-01211-0
Shannon, A., B. Selisko, N.T.T. Le, J. Huchting, F. Touret, G. Piorkowski, V. Fattorini, F. Ferron, E. Decroly, C. Meier, B. Coutard, O. Peersen & B. Canard. 2020. Rapid incorporation of Favipiravir by the fast and permissive viral RNA polymerase complex results in SARS-CoV-2 lethal mutagenesis. Nature communications (2020) 11:4682.
Shi, Z. & Z. Hu. 2008. A review of studies on animal reservoirs of the SARS coronavirus. Virus research, 133 : 74-87.
Sirotkin, K. & D. Sirotkin. 2020. Might SARS‐CoV‐2 Have Arisen via Serial Passage through an Animal Host or Cell Culture? BioEssays 2020, 42, 2000091: 1-7.
Wacharapluesadee, S., C.W. Tan, P. Maneeorn, P. Duengkae, F. Zhu, Y. Joyjinda, T. Kaewpom, W.N. Chia, W. Ampoot, B. L. Lim, K. Worachotsueptrakun, V. C-W. Chen, N. Sirichan, C. Ruchisrisarod, A. Rodpan, K. Noradechanon, T. Phaichana, N. Jantarat, B. Thongnumchaima, C. Tu, G. Crameri, M. M. Stokes, T. Hemachudha & L-F Wang. 2021. Evidence for SARS-CoV-2 related coronaviruses circulating in bats and pangolins in Southeast Asia.Nature Communication,12, 972. https://doi.org/10.1038/s41467-021-21240-1
Wenzel J. 2020a. Le SRAS et le COVID-19 n’ont pas de lien avec les civettes et les pangolins. Bulletin de la Société Française de Systématique, 62 : 14-22.
Wenzel, J. 2020b. Origins of SARS‐CoV‐1 and SARS‐CoV‐2 are often poorly explored in leading publications. Cladistics, 36(4) : 374-379.
World Organisation for Animal Health. 2020. COVID-19 Portal: Events in Animals. www.oie.int/en/scientific-expertise/specific-information-and-recommendations/questionsand-answers-on-2019novel-coronavirus/ events-in-animals/.
Zhan, S. H., D. E. Deverman & Y. A. Chan. 2020. SARS-CoV-2 is well adapted for humans. What does this mean for re-emergence? bioRxiv https://doi.org/10.1101/2020.05.01.073262
Zhou, P., X-L. Yang, X-G. Wang, B. Hu, L. Zhang, W. Zhang, H-R. Si, Y. Zhu, B. Li, C-L. Huang, H-D. Chen, J. Chen, Y. Luo, H. Guo, R-D. Jiang, M-Q. Liu, Y. Chen, X-R. Shen, X. Wang, X-S. Zheng, K. Zhao, Q-J. Chen, F. Deng, L-L. Liu, B. Yan, F-X. Zhan, Y-Y. Wang, G-F. Xiao & Z-L Shi. 2020. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature, 579 (7798) : 270-273.
Zhou, P. & Z. Shi. 2021. SARS-CoV-2 spillover events. Science, 371 (6525) : 120-122.
En ce cinquième anniversaire du confinement en France, cet article propose une synthèse des données scientifiques et autres révélations postérieures à juillet 2021 sur l’origine et les répercussions du Covid 19, faisant suite aux précédents articles écrits avant cette date par les mêmes auteurs.
Cet article vient compléter les 3 articles déjà parus dans Parlons Science. Il fait un état des lieux des variants et des vaccins disponibles à ce jour et se penche sur l’origine du virus SARS-CoV-2 qu’il parait indispensable d’élucider afin d’éviter de futures crises sanitaires mondiales.
Le virus SARS-CoV-2 s’est répandu très rapidement sur notre planète depuis la fin de l’année 2019. Responsable des symptômes associés à la maladie appelée Covid-19, ce virus a complètement bouleversé nos vies et la plupart de nos sociétés.
Un nouveau virus mortel vient de faire son apparition en Chine. A ce jour, plus de 2 000 personnes ont été infectées depuis le mois de décembre et 80 en sont mortes. 3 cas ont été identifiés en France. Quel est ce virus, d’où provient-il et comment se transmet-il ?
Votre abonnement à la lettre d’information : Muséum de Toulouse