Articlé rédigé parVALÉRIE MILS, Maître de conférence en biologie cellulaire – Faculté des Sciences et Ingénierie – Université Paul Sabatier à Toulouse
Cet article est le second de la série « DES VIRUS XXL QUI BOULEVERSENT LES DOGMES DE LA VIROLOGIE ». Le premier article a dévoilé l’existence de virus de taille exceptionnelle possédant une capacité codante parfois supérieure à certaines bactéries. Dans cette seconde partie, nous révélons d’autres propriétés originales de ces virus hors norme et évoquons les différentes hypothèses concernant leur origine évolutive.
Des parasites… de plus en plus autonomes !
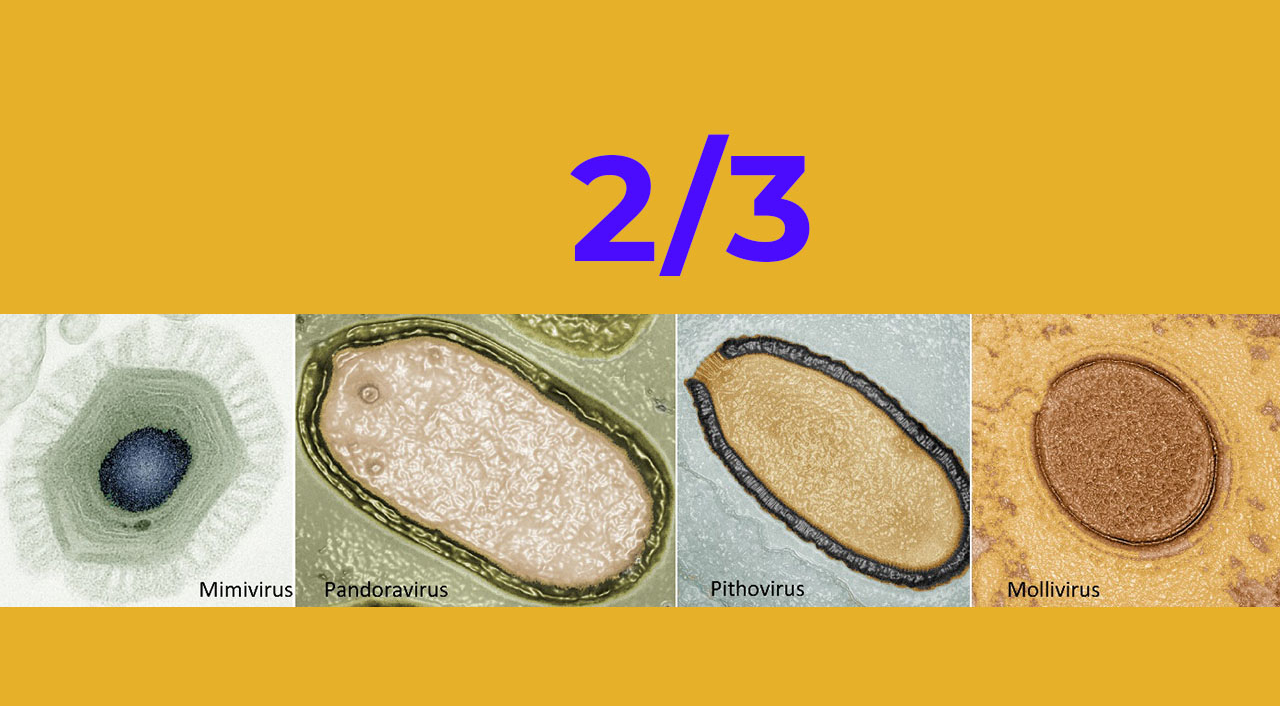
Les virus géants répertoriés à ce jour sont répartis en cinq sous-familles d’origines distinctes : les Mimiviridae, les Pithoviridae, les Pandoraviridae, les Mollivirus et les Phycodnaviridae.
Ces virus géants qu’on appelle également Girus, ont été rattachés au phyllum des Nucleocytoviricota plus connus sous l’acronyme NCLDV, pour Nucleo-Cytoplasmique Large DNA Viridae. En effet, ils possèdent tous un génome fait d’ADN double brin, circulaire ou linaire selon les familles de virus. Tous ces virus se distinguent des virus classiques par une taille exceptionnelle allant de 0,2 à 2µm (soit le double de la taille d’une bactérie ordinaire) et expriment un nombre incroyable de protéines, de 470 à 2500 protéines selon les représentants. Et ce n’est pas seulement leur taille et le nombre de leurs protéines qui font de ces virus des entités à part. Ainsi, lorsqu’on s’est intéressé aux nombreuses protéines produites par ces virus, on s’est rapidement rendu compte que la plupart d’entre elles étaient totalement inconnues (jusqu’à 90% dans le cas des Pandoravirus !). Ces protéines sont codées par des gènes dits « orphelins » c’est-à-dire qu’ils n’ont aucun équivalent connu dans l’ensemble des banques de données génétiques établies à ce jour. On ignore donc le rôle de ces protéines et l’origine des gènes qui les encodent… Qu’en est-il des autres protéines ? Là encore, les virus géants créent la surprise. Parmi les protéines connues qu’ils produisent certaines correspondent à des activités qui sont normalement l’attribut des bactéries, des archées et des eucaryotes c’est-à-dire d’organismes cellulaires et non de virus. Et cela explique l’originalité de leur cycle infectieux…
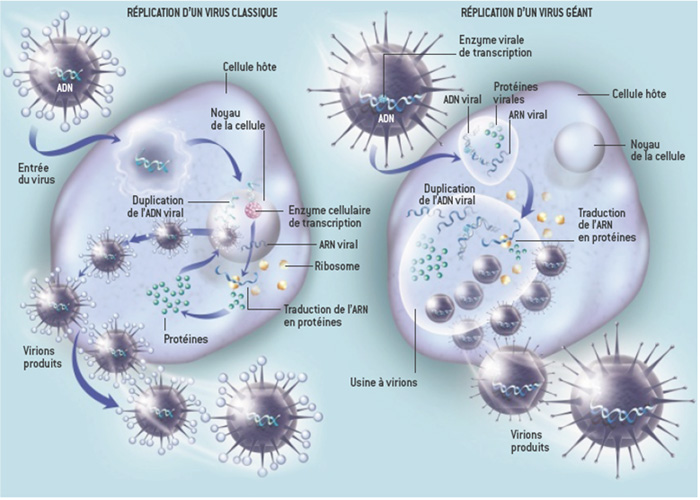
Modes de réplication du virus classique et d’un virus géant. Crédit : Claverie JM, Abergel C. Laboratoire IGS Université Aix-Marseille. Tiré de la revue Pour la Science n°415, mai 2012
En effet, contrairement aux virus à ADN classiques qui doivent importer leur génome dans le noyau cellulaire pour démarrer le cycle infectieux, certains virus géants comme les Mimivirus et les Pithovirus peuvent réaliser l’intégralité de leur cycle sans investir le noyau de la cellule hôte. Quelques heures après l’infection, ces virus induisent la formation d’une structure semblable à un « second noyau » nommée usine virale qui occupe une grande partie du cytoplasme alors que le « vrai noyau » montre des signes de dégénérescence. Au sein de ces « usines », le génome viral est répliqué et transcrit grâce aux ADN et ARN polymérases, hélicases et topoisomérases propres au virus. Plus surprenant encore, ces mêmes virus peuvent produire une partie de la machinerie de traduction des protéines ce qui est tout à fait inhabituel pour des virus ! Bien sûr, les virus géants ne sont pas totalement autonomes dans ces différentes fonctions : des facteurs doivent être importés du noyau vers l’usine virale pour compléter les machineries de réplication/transcription et la traduction de leurs protéines ne peut se faire en l’absence des ribosomes cellulaires. Mais quand même ! voilà des virus complexes qui brouillent sérieusement les limites entre virus et cellule c’est-à-dire entre vivant et non-vivant.
Les virophages et le système crispr-cas
En 2008, à l’occasion de l’identification d’un nouveau membre de la famille de Mimiviridae, le Mamavirus, on découvre d’autres virus beaucoup plus petits, co-infectant la cellule hôte. Ce petit virus pirate l’usine virale pour assurer sa propre propagation et cela au dépend du Mamavirus qui se met à produire des particules virales anormales. On venait donc d’identifier le premier parasite de virus géant, un virus de virus ! Ce premier représentant fut baptisé « Spoutnik », en référence au satellite artificiel du même nom. Depuis d’autres représentants furent identifiés chez les Mimiviridae : mavirus, zamilon, guarani… On en connait aujourd’hui environ une vingtaine. Par analogie avec les phages qui sont des virus de bactéries, ces virus de virus sont désignés sous le terme de virophages. Ils ne doivent pas être confondus avec de ce qu’on appelle les virus satellites qui ne peuvent se développer dans une cellule hôte qu’en présence d’un autre virus appelé virus helper. Les virus satellites n’affectent pas le cycle infectieux de leur virus helper, au contraire des virophages qui eux provoquent chez le Girus qu’ils parasitent la formation de virions anormaux défectueux.
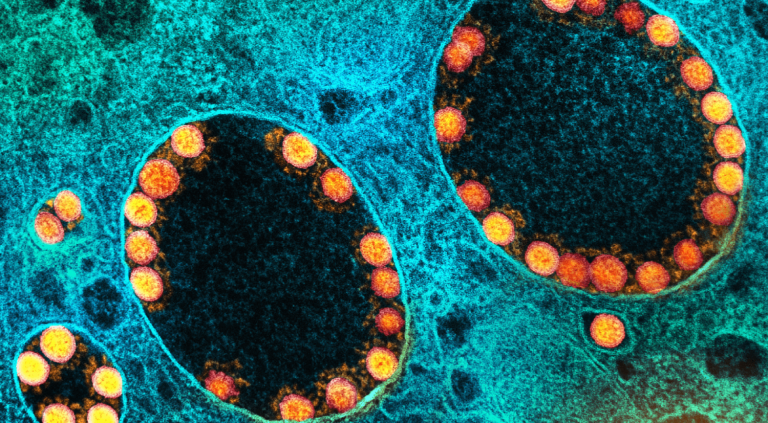
Image de microscopie électronique représentant les capsides du virus géant CroV et de son virophage associé, le mavirus. Coloration négative EM avec l’aimable autorisation de U. Mersdorf, MPI pour la recherche médicale, Allemagne. CC BY-SA 4.0 Sarah Duponchel et Matthias G. Fischer
Récemment, le génome entier de Mavirus, un virophage spécifique du Girus CroV, a été retrouvé sous forme intégrée dans le génome de l’amibe Acanthamoeba. Sous cette forme latente, Mavirus ne s’exprime pas. Par contre, il peut rapidement s’extraire de l’ADN et se réactiver en réponse à une infection par CroV. Ainsi, en hébergeant un virophage, l’amibe se dote d’un système de défense anti-Girus pérenne. Alors, est-ce la mort annoncée des Mimiviridae ? Bien sûr que non ! On sait que les virus exercent une pression de sélection permanente sur leurs hôtes et ceux-ci en exercent une en retour via le développement permanent de nouveaux moyens de défense. C’est la course aux armements auxquels se livrent, depuis qu’ils existent, parasites et parasités ! En accord avec ce principe, les Mimivirus ont bien trouvé le moyen de se préserver et leur réponse face aux virophages n’est pas sans nous surprendre de nouveau. En 2016, l’équipe de J.M. Claverie identifie dans le génome des Mimivirus des séquences correspondant à des morceaux de l’ADN du virophage Zamilon. Ces séquences sont bordées par des gènes codants pour des hélicases et des nucléases (qui déroulent et dégradent l’ADN). Si vous avez suivi les actualités autour des techniques d’édition du génome qui ont fait couler beaucoup d’encre dans les années 2010, vous aurez peut-être reconnu le dispositif de type CRISPR-Cas. Ce système a été identifié il y a longtemps comme le système de défense utilisé par les cellules bactériennes contre les phages et c’est la première fois qu’on montre qu’un virus utilise un système analogue pour lutter contre ses virophages ! Encore un caractère qui ne pourra plus être considéré comme propre aux organismes cellulaires ! Reste à savoir si c’est le virus qui a « emprunté » ce système aux bactéries ou…l’inverse !
D’où proviennent les virus géants et quel a été leur rôle dans l’évolution ?
Origine proto-cellulaire des virus géants
Vous l’avez compris, ces virus géants occupent une place à part dans le monde de la microbiologie. Pour cette raison, nombre de spécialistes tentent de déterminer leur origine et de comprendre la façon dont ils ont évolué. La plupart s’interrogent également sur leur statut : doit-on les considérer comme des organismes vivants ou des particules inertes ? Ce débat, qui s’étend d’ailleurs à tous les virus, est loin d’être clos mais voyons quelles sont les hypothèses débattues.
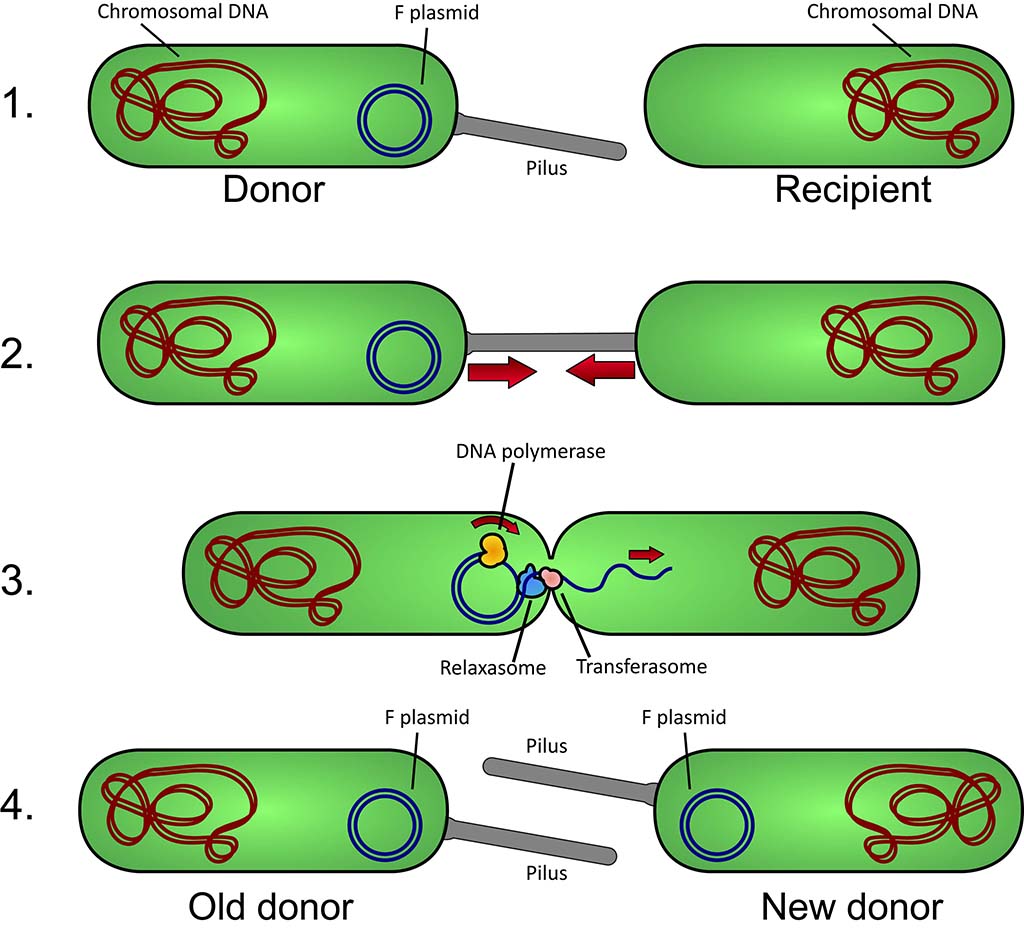
Pour introduire ce qui va suivre, rappelons quel est le scénario le plus probable de l’apparition de la vie. Tout d’abord, il faut abandonner notre vision binaire du vivant. La vie n’est pas apparue d’un seul coup grâce à l’accumulation séquentielle de fonctions spécifiques. Il faut plutôt imaginer un ensemble de systèmes moléculaires primitifs, issus d’environnements différents, chacun ayant acquis certaines compétences propres au vivant, mais pas toutes. Certains de ces systèmes appelés progénotes seraient dotés de capacité d’autoréplication, d’autres de compartimentalisation, d’autres enfin auraient développé un proto-métabolisme. Ces compétences ont ensuite été mutualisées entre les différents progénotes essentiellement grâce à des échanges de matériel génétique.
Schéma de conjugaison bactérienne. CC BY-SA 3.0 Adenosine, via Wikimedia
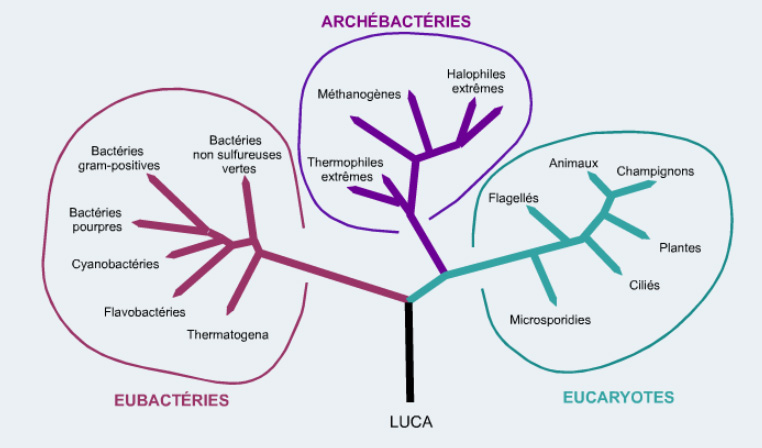
Cette coopération des progénotes a sans doute permis l’apparition d’ancêtres cellulaires (proto-cellules) qui ont ensuite co-évolués par coopération ou compétition et sous pression de sélection darwinienne. Au final, ces processus ont fait émerger une cellule, l’ancêtre commun de toutes les cellules actuelles appelé LUCA (Last Common Universal Ancestor). LUCA est donc déjà un système vivant autonome et complexe, adapté à son environnement et qui a évolué dans trois directions alimentant les trois grands règnes du vivant, bactéries, archées et eucaryotes.
Arbre phylogénétique des êtres vivants établi à partir de comparaison des ARN ribosomaux. LUCA est le dernier ancêtre commun de tous les êtres vivants. Crédit : Gérard et Nadine CIAVATTI via le site Web Evolution biologique
Mais revenons à nos virus géants. Quelle place ou quel rôle leur donner dans cette histoire des origines ? En absence de fossiles physiques, la seule façon d’explorer la généalogie des virus est de procéder à des analyses de phylogénie moléculaire. Lors de ces analyses, le degré de parenté entre organismes est corrélé au nombre de séquences génétiques ou de protéines qu’ils partagent. Dans le cas de virus géants, 60 à 90% des gènes codent pour des protéines totalement inconnues et n’ayant aucune ressemblance avec les protéines exprimées chez les bactéries, les archées et les eucaryotes. Il est donc peu probable qu’ils se soient construits sur la base d’un génome simple par l’accrétion de gènes «dérobés» aux organismes cellulaires. Si tel était le cas, on ne devrait pas y trouver autant de gènes orphelins dont on ignore toujours d’où ils proviennent. De plus, le mode de vie parasitique conduit généralement à la situation inverse : c’est le génome étranger qui est progressivement transféré vers le génome cellulaire ce qui conduit le parasite à une dépendance croissante vis-à-vis de son hôte. Nous avons donc une double énigme à résoudre : d’où proviennent tous ces gènes inconnus et comment expliquer la présence en petite quantité de gènes de bactéries, d’archées et d’eucaryotes ? Selon l’hypothèse la plus plausible, les virus géants seraient les descendants de progénotes ayant co-existé avec les ancêtres de LUCA. La sélection darwinienne n’aurait pas totalement éliminé ces progénotes qui auraient survécu en devenant des parasites obligatoires. Ce modèle explique l’absence d’homologues cellulaires pour de nombreux gènes viraux : ils correspondraient à des gènes n’ayant pas encore été transférés vers le génome de leur hôte. On peut s’interroger sur les conséquences pour la cellule hôte de ce transfert de gènes viraux (appelé transfert latéral ou horizontal de gènes). Il s’avère que ce processus a joué et joue encore un rôle essentiel dans l’évolution des organismes cellulaires. En effet, les génomes viraux seraient une source continue de nouveaux gènes. Ils se formeraient à partir de courtes séquences inter-géniques non codantes, appelées « proto-gènes », d’abord exprimés de façon aléatoire. A l’issue d’une sélection darwinienne, seuls les proto-gènes permettant l’expression d’une protéine phénotypiquement avantageuse sont stabilisés et transformés en gènes (voir Parlons science placenta). Ce phénomène est favorisé chez les virus géants, en particulier les Pandoravirus, par un grand nombre de séquences non codantes ainsi qu’une fréquence élevée de mutations et l’absence d’enzyme de réparation de l’ADN, ce qui permet de générer de nouveaux cadres de lecture sur l’ADN. Ce qui est excitant c’est que la caractérisation à venir de ces gènes et des protéines qu’ils codent pourrait conduire à la découverte de mécanismes ou de voies métaboliques originales n’ayant pas d’équivalent dans le monde cellulaire actuel. Si on en doutait encore, cela confirme le rôle puissant des virus dans l’évolution des génomes cellulaires ! Alors que penser maintenant de la poignée de gènes communs aux cellules (bactéries et eucaryotes) et virus géants: s’agit-il de gènes ancestraux mis en commun entre progénotes par transfert latéral de gènes avant l’émergence de LUCA ? Ces transferts ont-ils eu lieu plus tard entre cellule et virus et dans quel sens ? La question reste ouverte.
L’eucaryogénèse virale
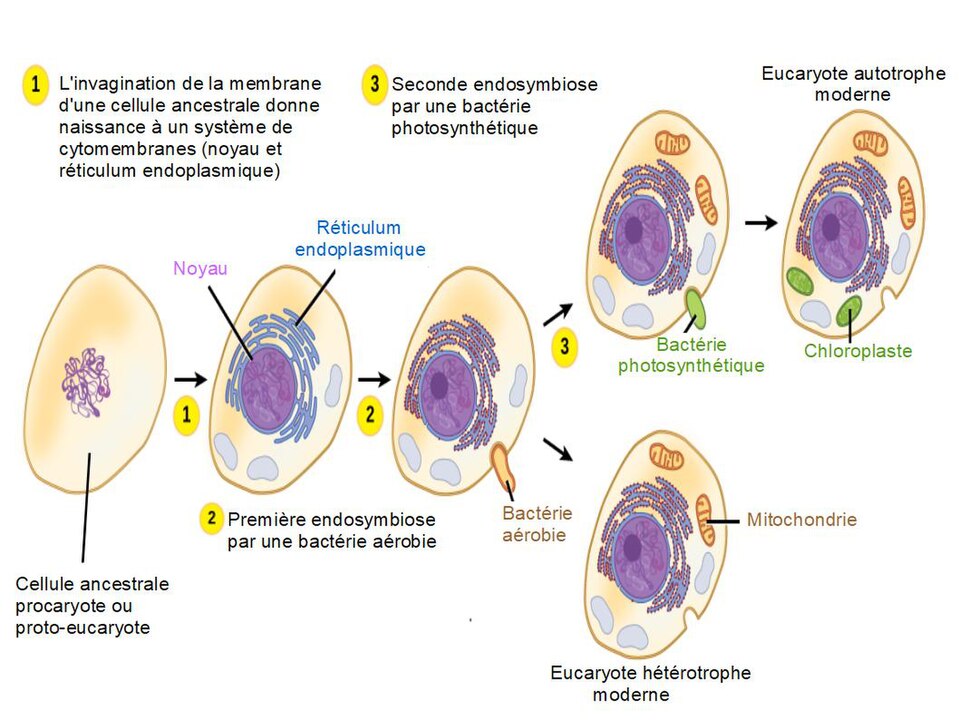
Une autre hypothèse émise il y a maintenant une vingtaine d’années confère aux virus à ADN (qui incluent les virus géants) un rôle central dans l’évolution de LUCA et l’émergence des eucaryotes. Les eucaryotes se distinguent des bactéries et des archées par la présence des mitochondries et par l’isolement de l’ADN dans un compartiment appelé noyau. Les travaux déjà anciens de Lynn Margulis ont permis de montrer que les mitochondries sont issues d’une endo-symbiose entre une archée et une protobactérie.
Origine endosymbiotique de la mitochondrie et du chloroplaste par un schéma de formation d’une cellule eucaryote hétérotrophe et autotrophe par absorption de bactéries. CC BY-SA 4.0 Salsero35, via Wikimedia
Cette bactérie endo-symbiotique a transféré la quasi-totalité de son génome vers le génome cellulaire pour ne conserver que sa fonction respiratoire capable de produire et mettre en réserve de l’énergie en synthétisant de l’ATP. Avant cela, un autre phénomène, probablement piloté par des virus à ADN a permis de créer le noyau. Cette hypothèse, qui ne pourra malheureusement jamais être démontrée, correspond au modèle de l’eucaryogénèse virale. En effet, l’usine virale qui se constitue dans la cellule hôte au moment de l’infection correspond au regroupement physique et fonctionnel du génome et des machineries de réplication et de transcription. Ce cluster exclut tous les organites cellulaires ainsi que la machinerie de traduction. La compartimentalisation physique de l’usine virale sous la forme d’un véritable noyau serait possible grâce au recrutement, en périphérie de l’usine virale, de membranes issues des autres organites cellulaires. Certains virus sont même capables d’induire la formation de novo d’une véritable enveloppe nucléaire autour de l’usine virale. Des pores situés à la surface de cette enveloppe favorisent la communication entre usine virale et cytoplasme, permettant, entre autres, l’export des ARN messagers vers la machinerie de traduction. Derrière la barrière de l’enveloppe, le génome viral est à l’abri des enzymes cellulaires susceptibles de le dégrader. Cette forte ressemblance entre l’usine virale et le noyau eucaryote suggère que ce dernier serait dérivé du premier.
On peut en effet imaginer que les gènes viraux contrôlant la formation de l’usine virale et de son enveloppe aient été détournés par une cellule ancestrale sans noyau pour constituer sa propre « usine à chromosomes ». De façon ironique et contre-intuitive, c’est l’accrétion par la cellule de gènes d’origine virale qui lui aurait permis de gagner ses caractéristiques de cellule eucaryote !
Et autres….
Le détournement des gènes viraux pour la formation du noyau eucaryote montre que, en dépit des apparences, l’infection par les virus ne correspond pas complètement à ce qu’on pourrait imaginer d’un strict parasitisme. D’autres gènes ont pu être détournés dans d’autres objectifs. Certains auteurs pensent en effet que les cellules ont pu apprendre des virus comment manipuler leur propre génome de façon à sélectionner des profils de gènes nécessaires à l’expression sélective de certaines fonctions. C’est peut-être grâce à cela que les processus de différenciation cellulaire propres aux eucaryotes pluricellulaires ont pu se mettre en place.
Notes
Cycle de virus à ADN Classiquement le génome des virus à ADN doit, suite à l’infection, être adressé au noyau de la cellule hôte pour être répliqué par une ADN polymérase de sorte à générer de nombreuses copies du génome viral. Les quelques gènes de ces génomes sont simultanément transcrits en leurs ARN messagers qui sortent du noyau et sont traduits en protéines virales grâce aux enzymes de traduction et aux ribosomes du cytoplasme. Les génomes et protéines virales s’assemblent ensuite pour former les virions qui lorsqu’ils atteignent un nombre élevé provoquent l’explosion (lyse) et donc la mort de la cellule hôte. Les virions peuvent alors partir à la conquête de nouvelles cellules à infecter.
En ce cinquième anniversaire du confinement en France, cet article propose une synthèse des données scientifiques et autres révélations postérieures à juillet 2021 sur l’origine et les répercussions du Covid 19, faisant suite aux précédents articles écrits avant cette date par les mêmes auteurs.